Экспертные обоснования прижизненной декапитации при рельсовой и других травмах (Экспериментальное исследование)
/ Ковалевский Г.В. // Судебно-медицинская экспертиза. — М., 1962 — №4. — С. 6-13.
")
Лаборатория патоморфологии (зав. — доцент Ю.Г. Целлариус) Института экспериментальной биологии и медицины Сибирского отделения Академии наук СССР
Kovalevsky, G. V.: Foundations for Expert Judgement of Intra Vital Decapitation in Rail and Other Traumas Experimental research)
Поступила в редакцию 8/V 1962 г.
В судебномедицинской практике при железнодорожной травме возникает вопрос, было ли отчленение головы от туловища прижизненным или посмертным. В этих случаях эксперт обычно дает заключение на основании совокупности данных следствия и вскрытия трупа.
Наше исследование посвящено поискам косвенных признаков, свидетельствующих о прижизненности повреждений, на основании изучения органов и тканей подопытных животных при отделении головы от туловища.
Материал и методика. Изучались органы и ткани гильотинированных кроликов и кошек. Эти опыты мы провели как вспомогательные при изучении патоморфологии острой смертельной кровопотери, которой в данном сообщении мы касаться не будем. Опыты состояли из следующих серий: 1) гильотинированные интактные кролики (10 опытов); 2) кролики, гильотинированные при максимальной глубине ганглионарной блокады, вызванной гексонием (10 опытов); 3) кролики, гильотинированные на высоте действия адреналина (12 опытов); 4) кролики, гильотинированные после введения ацетилхолина и антихолинэстеразных препаратов (6 опытов); 5) кролики, забитые супралетальными дозами адреналина (5 опытов); 6) кролики, забитые супралетальными дозами ацетилхолина (5 опытов); 7) гильотинированные кошки (10 опытов); 8) кошки, гильотинированные на фоне действия антихолинэстеразных препаратов (15 опытов).
Таким образом, во второй, третьей, четвертой и восьмой серии опытов перед обезглавливанием животным вводили фармакологические вещества, воздействующие на вегетативную нервную систему. Мы применили их для уточнения характера рефлекторных реакций, с которыми априорно должны быть связаны некоторые морфологические изменения при смерти животных от обезглавливания. На высоте действия названных препаратов животных обезглавливали и трупы их вскрывали через 5—15 минут после падения артериального давления до нуля.
В контрольных опытах (пятая и шестая серия) животные были забиты однократным внутривенным введением ацетилхолина (10 мг/кг) и адреналина (5 мг/кг). В качестве контрольных служили кролики, погибшие от пневмонии и кокцидоза.
Морфологическое исследование проводили с учетом кровенаполнения и кровераспределения в органах и тканях, которые изучали макроскопически и микроскопически с помощью бензидиновой методики Слонимского—Кунге и собственной методики избирательной окраски эритроцитов эритрозином в толстых срезах. Были использованы следующие гистологические и гистохимические методики: окраска гематоксилин-эозином, фукселин-пикрофуксином, импрегнация серебром по Гомори, окраска головного мозга по Нисслю, окраска на жир Суданом III и черным Суданом, окраска на гликоген кармином по Бесту, ШИК-реакция, реакция тетразониевого сочетания на белки.
Результаты исследования и их обсуждение. Продолжительность кровообращения у животных после обезглавливания различна. Наиболее короткой она была у кроликов, гильотинированных на фоне действия адреналина; у них артериальное давление достигало нуля в течение 30—90 секунд после обезглавливания. Наибольшая продолжительность кровообращения была у интактных гильотинированных животных, особенно у кошек, у которых падение артериального давления до нуля наблюдали на 3-й минуте и в начале 4-й минуты. Остановка сердца у обезглавленных животных отмечена значительно позже падения до нуля артериального давления. Нередко мощные сокращения сердца можно было видеть при вскрытии грудной клетки на 10—15-й минуте после обезглавливания. Типичных дыхательных движений после обезглавливания мы не наблюдали. У подавляющего большинства животных отмечены тонические судороги. Самые ранние судороги появлялись в середине 1-й минуты, а последний приступ судорог — нередко на 5-й минуте после обезглавливания. Из сосудов каудальной культи шеи кровотечение практически прекращалось к концу 1-й минуты. Кровопотеря при обезглавливании достигает Уз общего количества крови, которое нами принято за 6% веса тела.
Сущность общих морфологических изменений при обезглавливании сводится к малокровию органов и тканей, спастической реакции сосудов на кровотечение, явлениям перераспределительного лейкоцитоза и отмешиванию форменных элементов крови от плазмы. Эти изменения содержимого сосудов в настоящее время объясняют внезапными изменениями гемодинамики вследствие реакции сосудов на профузное кровотечение (Г.И. Мчедлишвили, 1958; Дибкэр, 1959) и возникающим в этих условиях перераспределением форменных элементов крови. Поскольку скорость циркуляции эритроцитов превосходит скорость циркуляции плазмы (Крейн и соавторы, 1959), то при кровотечении потеря эритроцитов должна преобладать над потерей плазмы.
При обезглавливании бывают характерные изменения органов: кровоизлияния под эндокард левого желудочка, своеобразная картина перераспределения крови в почках, аспирация крови в легких.
Аспирация крови в легких обусловлена инспираторной установкой грудной клетки во время судорог, поскольку дыхательные движения после обезглавливания, по-видимому, отсутствуют (записи дыхания мы не проводили). При отсутствии судорог у животных мы не наблюдали и крови в альвеолах, хотя отмечено ее попадание в трахею и главные бронхи. Легкие макроскопически выглядят различно в зависимости от наличия или отсутствия аспирации. В последнем случае легкие бывают или нормальной воздушности и удовлетворительного кровенаполнения, или же полуспавшиеся с очагами ателектазов и дистелектазов, которые выделяются относительно большим кровенаполнением; на этом фоне нередко можно наблюдать острую краевую эмфизему.
При наличии небольшой аспирации крови мы наблюдали умеренно выраженную тотальную острую эмфизему, а дистелектазы и ателектазы почти не встречали. При массивной тотальной и субтотальной аспирации крови легкие резко увеличены в объеме, несколько уплотнены, но тестоваты на ощупь. В случаях очень массивной тотальной аспирации культи почти вся кровь, обычно теряющаяся при обезглавливании, аспирировалась в легкие, и наружное кровотечение было очень скудным.
Помимо ателектазов, дистелектазов и острой эмфиземы, степень выраженности которой в основном соответствовала величине аспирации крови, иногда в легких, в просветах бронхов при микроскопическом исследовании обнаруживали клетки слущенного бронхиального эпителия. Поскольку явления слущивания эпителия бронхов наблюдали преимущественно в сериях опытов, где животные были обезглавлены после введения ацетилхолина и антихолинэстеразных препаратов, мы присоединяемся к мнению авторов (М.И. Касьянов, 1954; Н.Г. Пайков, 1960), считающих эти явления прижизненным процессом, обусловленным спазмом бронхов.
В сердце у обезглавленных животных наблюдали кровоизлияния под эндокард левого желудочка — пятна Минакова. У гильотинированных животных субэндокардиальные кровоизлияния преимущественно точечные и одиночные (не более 3—5). Наиболее часто пятна Минакова располагались в области межжелудочковой перегородки (в 18 случаях) и на сосочковых мышцах (в 11 случаях), реже — под митральными клапанами и в других местах. При микроскопическом исследовании кровоизлияния обнаружены не только в субэндокардиальных слоях миокарда, но и между листками самого эндокарда. Характерно, что перегородка и сосочковые мышцы — места наиболее частой локализации пятен Минакова — более малокровны по сравнению с другими участками миокарда левого желудочка, причем в глубоких слоях малокровие выражено ярче, чем в субэндокардиальных.
П.А. Минаков (1902) происхождение субэндокардиальных кровоизлияний в левом желудочке при кровопотере объяснял понижением давления в левом желудочке во время диастолы и «подсасыванием» крови из более глубоких слоев миокарда под эндокард; при этом во время систолы в относительно полнокровных субэндокардиальных слоях миокарда возникают диапедезные кровоизлияния.
В настоящее время мы располагаем достоверными физиологическими данными (Меесман, 1958, и др. ) об участии в наполнении сердца, помимо vis a tergo, еще и vis a fronte — присасывающего действия желудочков. Различают систолическое присасывающее действие, обусловленное движениями створок клапанов, и диастолическое присасывающее действие. Последнее отмечают в ранний период диастолы перед фазой быстрого наполнения желудочков; оно соответствует увеличению градиента давления между предсердием и желудочком за счет падения артериального давления в желудочке вплоть до абсолютно отрицательного (до —5 мм). Диастолическое присасывание увеличивается с уменьшением остаточного объема крови в желудочках, в связи с чем отмечено нарастание присасывания при кровопотере, искусственном стенозировании митрального клапана и других причинах, вызывающих уменьшение притока крови к желудочкам.
Таким образом, выводы П.А. Минакова о механизме кровоизлияний под эндокард при кровопотере в известной мере подтверждены данными современной экспериментальной физиологии. В настоящее время полагают, что диастолическое присасывание обусловлено энергией эластических сил, мобилизуемых систолой и освобождающихся во время диастолы. В этом процессе важное значение приписывают ретикулиновому каркасу сердца (Хорт, 1960) и физико-химическим факторам сцепления между ретикулиновыми волокнами и сократительными элементами мышечных волокон (Лютембаше, 1959). Состояние последних регулируется катионами К+ и Са++ и находится под контролем блуждающего нерва. На связь субэндокардиальных кровоизлияний с состоянием перевозбуждения блуждающего нерва указывает ряд исследователей (В.П. Десятов, 1951; Берблингер, 1917; Штюбель, 1924, и др. ). Она обнаружена при феномене так называемого ускользания сердца из-под влияния блуждающего нерва (О.В. Ульянова, 1953).
Наши опыты также свидетельствуют о значении функционального состояния блуждающего нерва для возникновения пятен Минакова. Так, у животных, гильотинированных на фоне действия ацетилхолина и антихолинэстеразных препаратов, пятна Минакова наблюдались чаще, чем у интактных гильотинированных животных: у кроликов почти в 2 раза, у кошек более чем в 2 раза. Кровоизлияния под эндокард левого желудочка у кроликов получены нами и без кровопотери, при смертельном отравлении ацетилхолином. Однако следует подчеркнуть, что и предварительное возбуждение симпатического нерва адреналином ведет к более частому возникновению пятен Минакова по сравнению с гильотинированными интактными кроликами (примерно в 1, 5 раза). Кровоизлияния под эндокард левого желудочка возникали и без кровопотери, при отравлении интактных кроликов адреналином.
Исходя из опытов О.В. Ульяновой, наблюдавшей кровоизлияния под эндокард при «ускользании» сердца из-под влияния блуждающего нерва, можно полагать, что возникновение пятен Минакова при этом феномене связано не с раздражением блуждающего нерва, а с резким усилением сократительной деятельности миокарда после «ускользания» вследствие безраздельного господства симпатического нерва над сердцем. На значение состояния мышцы сердца и способности ее к мощной сократительной деятельности для возникновения пятен Минакова указывает и В.П. Десятов. Этим, в частности, и объясняется отсутствие пятен Минакова при кровопотере у лиц с патологически измененным миокардом и почти постоянное их наличие при смертельной кровопотере у животных, которые редко страдают поражениями миокарда и сердца (В.П. Десятов, 1951).
Следовательно, не само по себе возбуждение и перевозбуждение блуждающего нерва, а изменение вследствие этого функционального баланса вегетативной иннервации сердца в сторону преобладания тонуса симпатической нервной системы (М.Г. Удельнов, 1948, 1949, 1957), по-видимому, вызывает возникновение пятен Минакова. Возбуждение блуждающего нерва ведет к ослаблению силы и урежению сокращений сердца, что является неблагоприятным условием для возникновения пятен Минакова. Однако при перевозбуждении блуждающего нерва, когда уже почти остановившееся сердце вследствие «ускользания» из-под влияния блуждающего нерва вновь начинает часто и усиленно сокращаться, при наличии других благоприятных условий (уменьшение притока и наполнения желудочка и вследствие этого усиление диастолического присасывания) становится возможным возникновение пятен Минакова. Характерно, что в наших опытах в условиях глубокой ганглионарной блокады, когда нарушается передача импульсов как по парасимпатической, так и по симпатической нервной системе, пятна Минакова у гильотинированных животных не возникали.
Обнаружив вполне определенное значение исходного состояния вегетативной нервной системы для возникновения пятен Минакова, мы все же полагаем, что противопоставление вакатного механизма возникновения пятен Минакова механизму нейрогенному (Берблингер, 1917; Штюбель, 1924) не обосновано, поскольку вегетативная нервная система имеет непосредственное отношение к механической работе сердца и тем самым участвует в создании ваката в полостях желудочков при дефиците наполнения сердца.
К числу изменений, характерных для прижизненной декапитации, следует отнести и обнаруженное нами своеобразное перераспределение крови в почках, которое встречается не только у гильотинированных животных, но и в случаях острой смертельной кровопотери при определенном характере кровотечения (Г.В. Ковалевский, 1961). Суть изменений (рис. 1) сводится к избирательному обескровливанию коркового вещества почек вследствие спазма междольковых артерий и афферентных артериол клубочков на фоне абсолютного или относительного полнокровия всего мозгового вещества или только полоски его, пограничной с корой почек. Полнокровие данной зоны мозгового вещества, равно как и прилежащей к ней узкой полоски коркового вещества, обусловлено тем, что сосуды этой кортико-медуллярной области почек обладают особой реактивностью, благодаря которой они остаются раскрытыми и максимально включаются в кровоток при спазме сосудов коры почек. Такое противоположное состояние сосудов основной массы коркового вещества по сравнению с состоянием сосудов кортико-медуллярной области способствует переключению большой части крови, поступающей в почки, в сосуды мозгового вещества, вследствие чего возникает избирательная ишемия коры почек. Этот феномен впервые описан Труэта и сотрудниками (1947, 1948) и получил название шунта Труэта. Он не является патогномоничным для кровопотери и встречается при других патологических состояниях.
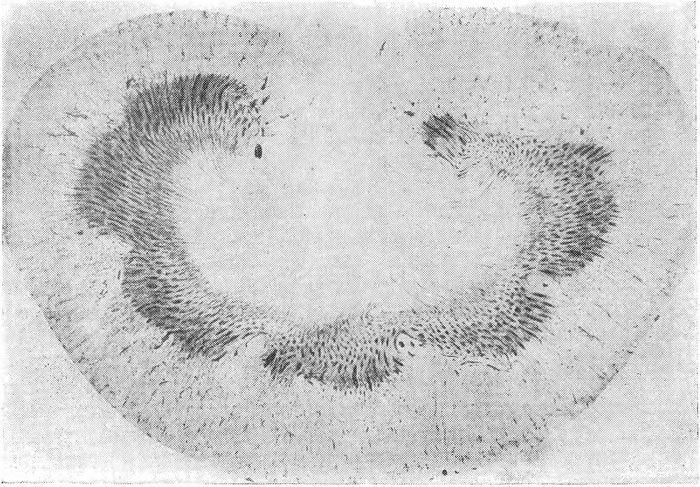
Рис. 1. Избирательное обескровливание коры почек (преимущественно средней зоны) на фоне полнокровия кортико-медуллярной области у гильотинированной кошки.
Окраска эритрозином. Целлоидиновый срез толщиной 100 µ. Лупа, увеличение 7×.
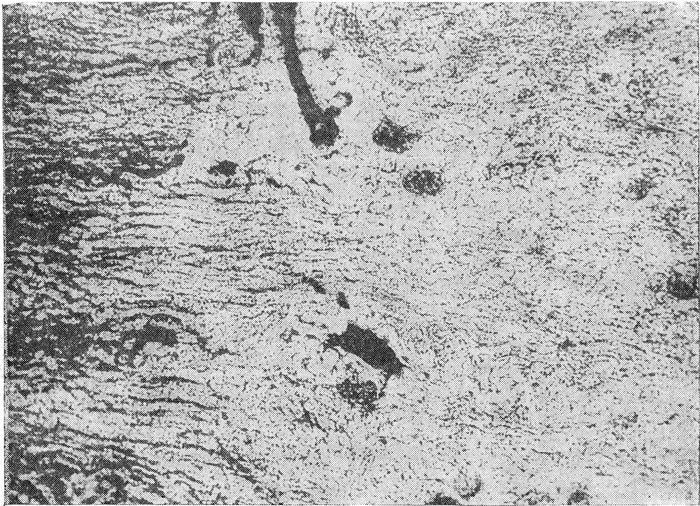
Рис. 2. Почка гильотинированного кролика. Обескровливание коркового вещества (выпадение рисунка клубочков) на фоне полнокровия пограничной зоны мозгового вещества (на снимке внизу видна только небольшая ее часть).
Окраска эритрозином. Толщина среза 100 µ. Ок. 7, об. 3, 7.
Характерно, что в наших опытах шунт Труэта отсутствовал у всех животных, гильотинированных на фоне глубокой ганглионарной блокады, а также у животных, обезглавленных на высоте действия адреналина. В первом случае не наблюдалось обескровливания коры почек ввиду отсутствия спастической реакции ее сосудов, во втором случае спастическая реакция была более резко выражена в сосудах кортико-медуллярной области, т. е. в состоянии резкого спазма находились сами пути переключения кровотока. Признаки шунта Труэта отсутствовали и в контрольном опыте при смертельном отравлении адреналином и ацетилхолином. У гильотинированных интактных кроликов характерное для шунта Труэта распределение крови в почках (рис. 2) встречалось в 5 из 10 опытов, у гильотинированных интактных кошек — в 9 из 10 опытов. В случае падения артериального давления до 0 ранее 2 минут после обезглавливания избирательного обескровливания коры почек не наблюдалось. Факт переключения кровотока почек в мозговое вещество при кровотечении у обезглавленных животных и неудачные попытки вызвать шунт почечного кровотока путем стимуляции сосудов адреналином и ацетилхолином свидетельствуют о том, что перераспределение крови в почках при кровотечении не зависит от центральных сосудистых рефлексов. Поэтому мы полагаем, что изменение кровотока почек по типу шунта Труэта относится к проявлениям ауторегуляции почечного кровотока. На возможность ауторегуляции почечного кровообращения благодаря самостоятельной реактивности сосудов почек, отличной от таковой других сосудистых областей, впервые указал В.М. Чернов (1949). Сейчас этот феномен интенсивно изучается.
В печени у гильотинированных животных наблюдаются дискомплексация ткани, нарушение радиарной структуры балок и диссоциация составляющих их печеночных клеток, вследствие чего малокровная ткань печени напоминает «булыжную мостовую». Исходя из тесной анатомической взаимосвязи между паренхимой печени и ее сосудистой системой, а также из экспериментальных данных о тесной зависимости пространственной структуры печеночной дольки от состояния гемодинамики в синусоидальной системе печени (Бароне и сотрудники, 1958, 1959), мы полагаем, что дискомплексация ткани печени является признаком внезапного падения общего и внутрипеченочного кровяного давления; в этих условиях нарушается опорная функция капиллярной системы печени по отношению к паренхиме и вследствие этого происходит диссоциация пространственной структуры балок печеночной дольки. Наличие в печени обезглавленных животных больших светлых как бы опустошенных (при окраске гематоксилин-эозином) печеночных клеток (рис. 3) не представляет собой какого-либо патологического явления, а является морфологическим отражением определенной функциональной стадии печеночных клеток и связано с наличием в них большого количества гликогена.
В заключение отметим, что при обезглавливании могут наблюдаться и изменения головного мозга, обусловленные непосредственным действием травмы. На месте расчленения шеи (на уровне I—III шейного позвонка) макроскопических кровоизлияний в спинной мозг мы почти не наблюдали, да и микроскопически они не всегда обнаруживались. Зато нередко встречались распространенные кровоизлияния в мягкие мозговые оболочки и субарахноидальные пространства в области продолговатого мозга, на основании мозжечка, моста и в большой цистерне. Хотя обезглавливание в наших опытах производилось одним ударом острого ножа, мы, однако, полагаем, что причиной такой контузии ствола головного мозга и мозжечка является гидродинамическое действие травмы («ликворный удар»).
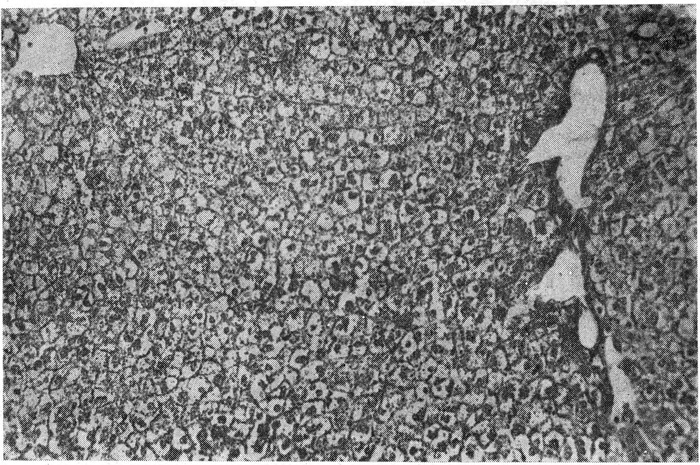
Рис. 3. Печень гильотинированного кролика. На фоне дискомплексации ткани среди клеток с зернистой «разреженной» цитоплазмой встречаются и «опустошенные» клетки, ШИК-реакцией и при окраске кармином по Бесту в них обнаруживается большое количество гликогена.
Окраска гематоксилин-эозином. Ок. 10, об. 10.
Выводы
- У обезглавленных кроликов и кошек наряду с повреждениями, непосредственно связанными с механической травмой, имеются общие изменения органов, обусловленные профузным кровотечением.
- В зависимости от интенсивности и продолжительности тонических судорог в легких на фоне острой эмфиземы наблюдается различной величины аспирация крови. Она обусловлена инспираторной установкой грудной клетки во время судорог после обезглавливания.
- В сердце обезглавленных животных встречаются пятна Минакова, в возникновении которых важное значение имеет исходное функциональное состояние вегетативной нервной системы.
- В почках у обезглавленных животных нередко обнаруживаются признаки так называемого шунта Труэта: избирательное обескровливание коры почек на фоне полнокровия кортико-медуллярной области, а иногда и всего мозгового вещества почек.
похожие статьи
Редкий случай травмы шеи при извлечении тела из воды / Виндгассен М., Ресслер Л., Тсокос М., Эстерхельвег Л. // Судебная медицина. — 2019. — №4. — С. 32-33.
Роль иммуногистохимического исследования в установлении прижизненности и выраженности огнестрельных повреждений мягких тканей / Богомолов Д.В., Федулова М.В., Шай А.Н., Павлова А.З., Збруева Ю.В. // Судебно-медицинская экспертиза. — М., 2018. — №6. — С. 46-47.
Морфологическая характеристика коры мозжечка при ожоговой травме / Морозов Ю.Е., Дорошева Ж.В., Горностаев Д.В., Колударова Е.М., Пиголкин Ю.И. // Судебно-медицинская экспертиза. — М., 2018. — №4. — С. 24-27.
Судебномедицинская характеристика смертельного травматизма на узкоколейных железных дорогах / Андрианов Л.П. // Судебно-медицинская экспертиза. — М., 1966. — №3. — С. 15-18.
О.Х. Поркшеян. Судебномедицинская экспертиза при железнодорожных происшествиях, 1965. / Огарков И.Ф. // Судебно-медицинская экспертиза. — М., 1966. — №2. — С. 54-55.
О продолжительности жизни и способности к действиям при смертельных повреждениях / Соколов Е.Я., Петрова А.В. // Судебно-медицинская экспертиза. — М., 1966. — №1. — С. 37.